Article for the “Bio/Mol/Text” competition: Recently, much attention has been paid to the development of cell replacement therapy methods, which are planned to be used for previously impossible treatment of injuries and diseases, including hereditary ones. Some potential therapies rely on the use of embryonic stem cells, which can differentiate into cells of any adult tissue. But the desired effect of stem cell treatment is not always achieved: people with paralysis of a limb or a few wrinkles on the face after therapy cannot even get out of bed and find bones in the eyelids. What makes cells go crazy? In this article, you will learn how complex epigenetic mechanisms not only ensure the successful differentiation of embryonic stem cells, but also sometimes cause them to turn into tumors.
Competition “Bio/Mol/Text”-2020/2021
This work was published in the “Free Topic” category of the “Bio/Mol/Text” competition 2020/2021.
The general partner of the competition is the annual biotechnology conference BiotechClub, organized by the international innovative biotechnology company BIOCAD.
The sponsor of the competition is SkyGen: a leading distributor of life science products on the Russian market.
Competition sponsor: the largest supplier of equipment, reagents and consumables for biological research and production.
"Book" sponsor of the competition - "Alpina Non-Fiction"
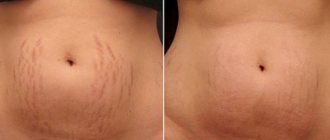
Preparation for the mesotherapy procedure
In order for the mesotherapy procedure to be most effective, during the course you need to adhere to several rules:
- Minimize skin exposure to ultraviolet radiation and use sunscreen with an SPF level of at least 40.
- Stop using blood thinners.
- Do not visit the sauna or bathhouse.
- Reduce physical activity. Before and after the session, it is better to give up fitness.
- Don't drink alcohol.
Whose, excuse me, beauty blog?
At what stage of embryo formation can embryonic stem cells (ESCs) be isolated? Here we will just whirlwind through the stages of early embryonic development, and details can be found in the article “How to make a neuron from a fibroblast?” [1]. After fertilization, the zygote divides by mitosis and on the fourth day it becomes a morula, on the fifth - a blastocyst, which consists of the inner cell mass (ICM) and trophectoderm, and on the seventh it is ready for implantation into the placenta [2] (Fig. 1).
Figure 1. Stages of preimplantation development of the human embryo. Phase-contrast images of human embryonic development from day (d) 0 to day 7. White arrow points to the ECM, gray arrow to the trophectoderm.
[2]
Figure 2. Hierarchy of stem cells. FSC - follicular stem cells, MSC - mesenchymal stem cells, HSC - hematopoietic stem cells, NSC - neural stem cells
[44]
ESCs are cells derived from the ECM. They can divide indefinitely and differentiate into all types of embryonic and adult cells. This ability is called pluripotency [3] (Fig. 2). The pluripotency of ESCs has given rise to great hopes that, when introduced into a patient’s body, they will be able to restore or replace the lost functions of an organ or tissue. This approach, called cell therapy , could be used to treat a variety of diseases, such as Parkinson's disease, diabetes and spinal cord injuries [3].
But using ESCs is unethical because they are obtained from human embryos, and it is also difficult because they are rejected by the patient’s body after transplantation [4]. To overcome these obstacles, K. Takahashi and S. Yamanaka generated induced pluripotent stem cells (iPSCs), which was published in Cell in 2006 [5]. To do this, genes for four transcription factors were introduced into somatic fibroblast cells: Oct4, Sox2, c-Myc, and Klf4. As a result, gene expression and the epigenetic pattern of the genome, which provides the phenotype of a differentiated cell, have changed.
They also want to use iPSCs for cell therapy, but so far their use is limited to the creation of cell lines-models for studying rare genetic diseases and testing the effectiveness of drugs against them. The production, application and limitations on the use of iPSCs are described in the article “Nobel Prize in Physiology or Medicine (2012): induced pluripotent stem cells” [6]. And in the article “There Was a Simple Cell, It Became a Stem Cell” [7], the experiment of Shinya Yamanaka and the functions of transcription factors with the help of which the Japanese scientist’s group reprogrammed somatic cells are described in more detail.
Many types of cells of a multicellular organism with the same set of genes are formed precisely because of differences in their expression, which depends on the epigenetic pattern of the genome. After each division of the mother cell, the daughter cells may or may not retain its epigenetic pattern. If, after division, the daughter cell differentiates, its epigenetic pattern changes, causing it to acquire a new phenotype [8]. Thus, through differentiation from a mass of identical ESCs, specialized cells of all tissues of our body are formed [3].
Gene structure and epigenetic modifications
The creation of RNA from a DNA template in a cell is controlled by DNA regulatory sequences located at the edges of genes: promoters, enhancers and silencers. Promoters are located near the gene and consist of a core promoter and a proximal (near) promoter. The main promoter is needed to initiate transcription, and the proximal promoter is needed to regulate the intensity of transcription. Enhancers and silencers are located at a distance of many thousand base pairs from the gene and regulate the activity of promoters [9] (Fig. 3).
Figure 3. Gene structure
[9]
Regulatory sequences allow a cell to increase, suppress, or even completely stop or activate the expression of its genes through special proteins—transcription factors (TFs) [9]. But control of expression is not at all the prerogative of silencers and promoters: a major role is also played by changes in the accessibility of chromatin for binding to TFs through epigenetic modifications (for details, see the article “Epigenetics: the invisible commander of the genome” [10]).
Epigenetic modifications include chromatin remodeling, post-translational modifications of histones, and methylation of DNA nucleotide bases. In particular, methylation of the promoter and enhancer in most cases suppresses gene expression, although any DNA region can be methylated [11].
Chromatin remodeling involves the removal and movement of nucleosome remodeling proteins, which consist of core histones and DNA wound on them and prevent the binding of other molecules to the polynucleotide. The distribution of nucleosomes in the genome is heterogeneous: they are densely located within heterochromatin, but there are few of them on regulatory sequences and transcribed genes [12] (Fig. 4). Post-translational modifications of histones (about which there is material in the article “Pills for the Epigenome” [13]) change not only the affinity of nucleosomes for chromatin remodelers, but also the affinity of histones for DNA [14].
Figure 4. Diagram of chromatin organization. The DNA wraps around the histone octamer to form a nucleosome. Chromatin can form loops held by cohesin or other proteins. The chain of nucleosomes is compacted into topologically associated chromatin domains that interact over long distances and form chromatin compartments that can be transcriptionally active (A) or inactive (B). The interphase chromosome consists of several compartments that together form the territory of the chromosome.
[14]
Core histones undergo acetylation, methylation, phosphorylation, ubiquitylation (about ubiquitin, read the article “Ubiquitin” [15]), sumoylation (for more about the modification with a strange name, see the article “SUMO: Japanese wrestling or a unique post-translational modification?” [16] ), deamination, etc. The most common types of histones are acetylation, methylation, and phosphorylation [8].
How it works? For example, lysine acetylation changes the charge of ANA from positive to zero, which likely weakens DNA-protein interactions and converts tightly packed chromatin into a molecule open to transcription. Histone acetylation is required for embryonic cell differentiation [8]. Methylation of lysine and arginine increases the positive charge and hydrophobicity of ABA [17]. The effect on gene expression depends on how many methyl groups are attached to the amino acid and which amino acid is methylated. The balance between methylation and acetylation in the enhancer and promoter regions of a gene determines its expression level [8].
Phosphorylation increases the overall negative charge of the histone tail, preventing interaction with DNA. In stem cells, histone phosphorylation is involved not only in chromatin remodeling, but also in the repair of damaged DNA during differentiation [8].
In addition to core histones, there are several other variants of linker histone H1. The timing of their expression in the cell and their affinity for chromatin appear to play an important role in development. Weaker binding of linker histones to chromatin contributes to the creation of totipotency and maintenance of pluripotency of early embryonic cells, as well as differentiation [18].
Epigenetic modifications are regulated by transcription factors. Some of them were used by K. Takahashi and S. Yamanaka to obtain iPSCs from fibroblasts [19].
Detailed description
The earliest cells in the human body are stem cells. They are formed immediately after conception in a fertilized egg. The ability to become any cell is their main distinguishing quality, the so-called pluripotency. As the embryo grows, the stem cells form its brain, liver, stomach, and heart. Even after birth, there are still a lot of them in the child’s body, but every year there are fewer of them; by the age of 20, a person has practically no stem cells. This is a scientifically proven fact. But an adult also needs these cells - they always replace those damaged in the event of illness of any organ. Throughout life, organs with diseases become much more numerous, but stem cells decrease, so a person ages.
Transcription factors
Transcription factor (TF) genes occupy from 3 to 10% of the eukaryotic genome. TFs influence gene transcription by attaching to DNA regulatory sequences via a DNA-binding domain. The DNA-binding domain recognizes the nucleotide sequence of the promoter, forming hydrogen bonds and Vander Waals contacts between it and the residues of its AMKs. Nucleotide bases are contained within a double helix, so most TFs bind to the major groove of DNA [20] (Fig. 5). The affinity of TFs for promoters is determined by interactions with the sugar-phosphate backbone, including positively charged amino acids with phosphates.
Figure 5. Illustration of the major and minor grooves using the example of siRNA (small interfering RNA).
adapted from [20]
Common TFs help RNA polymerase II bind to the main promoter, promote DNA unwinding, and the transition from transcription initiation to RNA chain elongation [9], [21]. TFs bind to proximal promoters and change the affinity of the main promoter for RNA polymerase. Thus, the transcription factor Oct4, which was used to reprogram somatic cells into iPSCs, binds to the promoters of its target genes and can both promote transcription and suppress it [2]. TF activators or TF repressors of transcription bind to enhancers and silencers [9]. Repressors and activators can compete for the same DNA target site or block each other's actions [22].
TFs interact directly or through other proteins using protein interaction domains, stabilizing the transcription complex and increasing the intensity of transcription [21], [22]. To maintain the pluripotency of ESCs, Oct4 also interacts with other TFs: Sox2, Sall4, Zfp143, etc., which we will discuss in more detail below [4]. TFs also bind to coregulator proteins, which can activate (coactivators) or suppress (corepressors) gene expression. Some of them unite all the molecules involved in transcription like a scaffold, some catalyze reactions of epigenetic modifications. It is known that Oct4 cannot do without the help of coregulators [4], [22], [23].
results
Those patients who underwent stem cell rejuvenation courses at research institutes noted that after just three weeks, fatigue disappeared, body tone increased, visual acuity appeared, wrinkles smoothed out a little, men experienced increased libido and improved potency. As you can see, the results of body revitalization therapy in cosmetic clinics and research institutes are the same, although their methods are completely different.
Research institutes use a special cell growth factor protein, and beauty salons use additional mesotherapy. All these additional injections and procedures that come along with stem cell injections, according to doctors, are aimed at insuring clinics against the lack of results of stem cell treatment, since mesotherapy and additional protein have long been known as an excellent and effective way to smooth out wrinkles.
Cell therapy specialists are silent about whether there were negative results or no result at all. And there are such cases, patients did not notice any changes even after 3-6 months, but neither the clinic nor the research institute reimburses the costs in any way, because they do not guarantee that the body will find the strength to recover.
Pluripotency factor Oct4
Over time, researchers came to the conclusion that the main regulators of transcription in pluripotent stem cells are the transcription factors Oct4, Sox2 and Nanog. Let's look at how these TFs cope with their task using Oct4 as an example [4].
Octamer-binding proteins (Oct) are a group of TFs that specifically bind to the 8-nucleotide sequence ATGCAAAT [24]. Oct4 activates the expression of genes that support pluripotency and self-renewal of SCs, and at the same time suppresses the expression of genes that promote differentiation and represses transcription factors specific to cells of specific tissues [4].
There is a group of targets that are both activated and suppressed depending on the level of Oct4 expression. But basically, this TF clearly acts as either an activator or an inhibitor of the transcription of its target genes [4].
Let's see what the most famous Oct4 target genes are.
Table 1. Functions of the described genes and their relationship with Oct4
| Characteristics | Gene name | ||||||||
| Utf1 | Fgf4 | Zfp206 | Osteopontin | miR-302 | Cdx2 | Stk40 | Hand1 | hCG | |
| Oct4 activates expression | + | + | + | + | — | — | — | — | — |
| Oct4 represses expression | — | — | — | — | — | + | + | + | + |
| Gene involved in differentiation | + | — | — | — | — | — | — | — | — |
| Gene involved in embryo development | — | + | — | — | — | — | — | — | — |
| Gene involved in maintaining pluripotency | — | — | + | — | + | — | — | — | — |
| The gene is involved in the formation of extraembryonic tissues | — | — | — | — | — | + | + | + | — |
| The gene is involved in trophoblast implantation into the uterus | — | — | — | — | — | — | — | — | + |
| Gene involved in cancer formation | + | + | — | + | — | — | — | — | — |
The role of Oct4 target genes in ESC differentiation
Oct4 activates the expression of Utf1, Fgf-4, Zfp206, osteopontin, miR-302 [4]:
- Utf1 (undifferentiated embryonic cell transcription factor 1) is involved in chromatin organization, maintaining the rate of cell division, and the formation of teratomas. ESCs with reduced levels of Utf1 fail to differentiate properly during exit from pluripotency [25], [26];
- Fgf-4 (fibroblast growth factor 4) was originally considered an oncogene. Expressed in certain tissues of the embryo, maintains the viability of the blastocyst, plays a role in the growth and formation of the developing limb [27];
- Zfp206 (zinc finger protein) is expressed during embryogenesis but is absent in all adult tissues except testes. Overexpression of Zfp206 makes ESCs resistant to differentiation [28];
- Osteopontin is involved in bone breakdown, immune responses, biomineralization and wound healing. Overexpressed in many cancers, it promotes migration, invasion and self-renewal of cancer cells [29], [30];
- miR-302 is a cluster of eight microRNAs. Specifically expressed in ESCs. A member of this cluster, miR-302a, inhibits the translation of cyclin D1 (a regulator of the G1 phase of cell division) to maintain the cell cycle characteristics of ESCs [4].
Oct4 inhibits the expression of Cdx2, Stk40, Hand1, hCG [4], [31]:
- Cdx2 (caudal type homeobox 2) is required for trophectoderm formation [4], [32];
- Stk40 is involved in the formation of extraembryonic endoderm in mice [4];
- Hand1 (heart and neural crest derivatives-expressed transcript 1) plays a role in cell differentiation and embryo development. Begins to be expressed during preimplantation development; after implantation, it is expressed only in placental trophoblast cells [32-34];
- Human chorionic gonadotropin (hCG) prepares the endometrium for implantation and promotes successful implantation of trophoblast into the uterus. Coordinates interactions between the blastocyst and uterine tissues, regulates angiogenesis and immunity [35].
Judging Oct4 by its effect on these 9 genes, some conclusions emerge. Oct4 suppresses the expression of genes whose products promote the formation of extraembryonic trophoblast tissue (Cdx2, Stk40, Hand1) and its implantation into the uterine wall (hCG). And genes activated by Oct4 are important for cell differentiation immediately after ESC exit from the pluripotent state (Utf1), embryo development (Fgf4), and maintenance of ESC pluripotency (Zfp206, miR-302) [36] (Fig. 6).
Figure 6. Oct4 expression in the early embryo.
adapted from [36]
Moreover, among the genes positively regulated by Oct4, there are those whose expression promotes the formation of cancer tumors (Utf1, Fgf4, osteopontin). Oct4 itself, which is not expressed in normal adult tissues, has been found in some human cancers. High expression of Oct4 promotes tumor development, metastasis, and increased cancer-related mortality. But in people whose tumors express moderate or low Oct4, this is not observed [37]. This can be explained by the fact that stem and cancer cells have some similarities, specifically the ability of cells to divide indefinitely, which Oct4 supports in ESCs [38].
How does it depend on age?
If a young body needs approximately 20-35 million cells to maintain tone, then 200 million may not be enough for a lady of pre-retirement age with a bunch of diseases. According to experts, such a high price is justified, because growing cells is a process that requires knowledge and high technology, and is therefore very expensive. If you are offered such procedures at a lower price, then most likely these drugs are not related to stem cells.
There are, however, state scientific institutes where injections are cheaper, but the price still starts from 5 thousand US dollars. They use bone marrow stem cells. Scientific institutes also use special cell growth factors – peptides. Since stem cells, when injected, cannot find a damaged organ, proteins show them the way, which turn on the work of the body’s cell, forcing it to work and look for means of self-healing.
Auto- and cross-regulation of transcription factors
The core list of Oct4 target genes is not limited to those listed above and consists of 33 genes that often encode transcription factors (as do the target genes of other pluripotency factors, Sox2 and Nanog). In order to cope with the regulation of such a large number of genes, Oct4 interacts with other TFs and coregulators [4]. Oct4, Sox2, and Nanog in ESCs together occupy many regulatory sequences of their target genes, play a major role in maintaining pluripotency, and activate each other’s expression [39]. For example, the regulatory sequences of many Oct4 target genes contain sites that can be bound by Sox2, separated from the Oct4 binding site by only a few nucleotides, suggesting that these TFs cooperate closely.
Oct4 also interacts with other TFs that support pluripotency (Sall4, Zfp143, Zfp206, Esrrb, Dax1 and Tcfcp2l1). Some of them (Esrrb, Tcfcp2l1 and Dax1) are helped by Oct4 to target common sites. Oct4 and pluripotency TFs create autoregulatory and cross-regulatory loops [4]. In autoregulation, TF binds to its own promoter and either activates or represses transcription. Autoregulation makes it possible to determine and control TF concentrations in the cell without intermediaries, which also need to be regulated. Cross-regulation is the regulation of the expression of one transcription factor by another [40], [41], (Figure 7).
Figure 7. Interaction of Oct4 with TF. ( A ) Autoregulation: the initial transcription activator ( HA ; initial transcription activator) initiates ( I ) expression of gene X (box with arrow), which maintains ( R ) its own expression by directly binding to its promoter. ( B ) Cross-regulation: activation of Y gene expression by X gene.
adapted from [41]
Due to the fact that the pluripotency factor Oct4 alone has more than three dozen target genes and a complex network of interactions with other transcription factors, predicting and, even more so, controlling the behavior of pluripotent cells transplanted into the patient’s body is a very ambitious and still open task. In addition, Oct4, Nanog and Sox2 are sometimes expressed, including in cancer cells, and therefore can contribute to the development of cancer [42]. This may be a major obstacle to the use of ESCs in cell therapy.
Stem cell storage
Umbilical cord blood stem cells, as well as biological material isolated after collecting the placenta and umbilical cord, are stored in the Cryobank. Here you can see how all the processes associated with the processing and storage of umbilical cord blood, placenta and umbilical cord occur in our Cryobank.
After preliminary preparation, the isolated stem cells are cryo-frozen and stored in special containers with liquid nitrogen for any length of time.
As soon as the owner needs them, Cryobank will issue them upon request and deliver them completely intact to any clinic where the biological product will be used. You can learn everything about storing umbilical cord blood stem cells on our website or from the consultants of the Cryobank Institute of Cell Therapy. stem cells
Cell therapy today
Despite the popularity of the topic of stem cell therapy, there are no cases of successful treatment of anyone with ESCs in the literature. It is not surprising, because the more differentiation pathways are open to cells, the more serious the side effects of their introduction into the patient’s body can be.
But even the consequences of a facelift using mesenchymal stem cells (MSCs) that are more differentiated than ESCs can be unexpectedly severe. A US resident experienced them herself after undergoing a procedure, the essence of which was to extract MSCs from the client’s adipose tissue and return them under the skin of the face, especially around the eyes. The cosmetology clinic clearly failed to cope with its task: the woman’s eyelid hung over her right eye, and the area around it was swollen. After 3 months, the American woman came to the doctors with a complaint that she could not open her eyes without severe pain and that every time she did raise her eyelid, a sharp sound was heard, similar to the click of tiny castanets. The woman went under the knife, and six and a half hours later, surgeons removed small pieces of bone from her eyelid and the tissue surrounding the eye - they were the ones who clicked due to friction against each other. How did it happen that the bones ended up in such an inappropriate place? The fact is that in addition to MSCs, cosmetologists also injected the woman with an anti-wrinkle dermal filler, the main component of which is calcium hydroxyapatite, which triggers bone differentiation of MSCs. Plastic surgeons did not take this into account, endangering the health of their client.
What can stem cells with greater potential do? The answer is provided by the case when a boy with ataxia-telangiectasia (a congenital neurodegenerative disease) in a Moscow hospital was injected into the brain and its membranes with neural fetal stem cells (more differentiated than embryonic ones, but not very far from them in their “power”) [43] . Four years after this operation, the patient was diagnosed with several foci of a brain tumor. Molecular and cytogenetic studies have shown that the tumor cells are different from the patient's cells, which suggests that it develops from transplanted SCs.
A similar story happened with American Jim Gass. The man wanted to fully rehabilitate himself after a stroke, due to which his left leg became weak and his left arm was paralyzed. For this, Jim sought treatment in clinics in various countries until he settled on Mexico, where he was injected with fetal stem cells delivered from Russia. However, after some time, the American was struck by paralysis of the entire body below the neck. Upon examination, doctors discovered a tumor in the lower part of his spine, which turned out to be a huge mass of bloody tissue from poorly differentiated foreign cells. Apparently, this was the reason for the deterioration of Gass’s condition. Doctors were only able to slow down the aggressive growth of the tumor, but not stop it.
Now imagine what will happen if you inject embryonic stem cells into a patient. Scary? Me too. Although there is no need to strain your imagination: injection of SC into laboratory animals is a standard method of testing them for pluripotency. The above-mentioned experiment by Shinya Yamanaka could not do without it.
Why do transplanted stem cells behave “inappropriately”? The reason is that they find themselves in an unusual environment. The division and differentiation of SCs are regulated by a complex system of interactions with the intercellular substance and neighboring cells, and when SCs are artificially introduced into the body, the necessary environment is often not created, and the cells are lost among unfamiliar signals, which can lead to aggravation of the patient’s condition.
There is only one type of stem cells that have been learned to be used completely safely in medicine - hematopoietic stem cells (HSCs), the progenitors of blood cells. If a patient has a mutation that impairs hematopoiesis, he can receive a donor's HSCs or his own HSCs. In the second case, cells are first removed and the mutation is corrected using genetic engineering.
Biomolecules’ special project “Gene Therapy” talks about the symbiosis of genetic engineering and medicine.
But how can diseases not related to hematopoiesis be treated if other SCs are unsafe to use? It is possible to treat deep wounds, restore cartilage, create a cornea and lens if you remove differentiated cells that are still capable of division, multiply them in the laboratory and return them back. If there are no cells in the body suitable for treating a particular disease, you can take SCs from the patient and create conditions in the laboratory under which they would differentiate into the necessary cells. In this way, researchers propose to obtain cells for the treatment of diabetes, Parkinson's disease, leukemia, and retinal restoration. However, SC in the adult body is usually insufficient for cell therapy. This problem could be solved by iPSCs, but the technology for their production is not yet effective enough: very few cells achieve pluripotency, and in animal experiments it has been shown that they also induce an immune response. This may be due to the fact that reprogramming provokes mutations.
Probably, due to the need to develop regenerative medicine, we will be forced to create more advanced technologies for obtaining pluripotent stem cells and controlling their behavior in the patient’s body. And here it is no longer possible to do without studying the mechanisms of maintaining pluripotency of ESCs, in which transcription factors, including Oct4, play an important role.
Umbilical Cord Blood Stem Cells: Benefits
The main feature of umbilical cord blood stem cells is that they can be autologous. Parents who preserve the cord blood (and often also the umbilical cord and placenta) of their newborn child provide lifelong insurance in the event of serious illness or injury. Every year, new discoveries in medicine are made around the world related to the possibilities of using stem cell biological products. Therefore, in the future, a child who has such “insurance” will receive medicine for many possible diseases. Since the isolated stem cells are autologous, there will be no immune conflict during transplantation and new cells will not be rejected by the body.
One of the advantages of collecting cord blood is that the process is completely safe and painless. Obtaining stem cells from this biological material, as well as from the umbilical cord and placenta, does not contradict any ethical standards.
Autologous stem cells can be used not only directly for the child from whose blood they are isolated. In some cases, they may also be suitable for treating his siblings, and sometimes one of the parents, because there is a possibility of immune similarity with the donor’s relatives.
From preserved stem cells it is also possible to obtain a biological product that is used for cosmetic purposes and in gerontology to rejuvenate the body.
So, the main advantages of collecting and storing umbilical cord blood and placenta cells are:
- Safety and accessibility of collection in any maternity hospital.
- Full immune similarity with further use.
- Possibility of using cages for close relatives.
- Ethical and humane: when isolating cells, the body does not suffer and there are no risks for it.